
Introduction
糖尿病主要包括Ⅰ型糖尿病、Ⅱ型糖尿病和妊娠期糖尿病。国际糖尿病联盟2021年的报告显示,全球有5.37亿成年人患有糖尿病,预计到2030年将增至6.43亿,2045年将增至7.83亿。2021年全球每10人中就有1人患糖尿病,2045年每8个成年人中就有1人患糖尿病,增加了46%。特别是非洲2021年大约有2400万成年人(20-79岁)患有糖尿病,预计到2045年将增加到5500万,增长134%(图1)。此外,2021年中国有1.4亿糖尿病患者,预计2045年将上升到1.74亿。值得注意的是,Ⅱ型糖尿病占糖尿病的90%以上。因此,预防或减缓Ⅱ型糖尿病的研究是一个热点话题。
Ⅱ型糖尿病是一种常见的慢性代谢性疾病。长期高血糖和代谢紊乱(糖、脂)可导致全身组织和器官损伤,如肝脏、肾脏、心血管和神经系统等,危及人们的生命健康,甚至影响其正常生活。Ⅱ型糖尿病的高发病率和增加的患病率带来了社会和经济负担,引起了全世界的关注。
降低患者血糖的常见策略暴露出诸多弊端。尽管二甲双胍是Ⅱ型糖尿病初期最受推崇的治疗方法,但是常发生胃肠道副作用。皮下注射胰岛素也是一种有效的策略,但注射部位的反应和低血糖的风险不容忽视。食物中的生物活性成分因其安全低毒的特性越来越受到研究者的青睐。特别是膳食多糖因其高安全性、低副作用和潜在的糖代谢调节作用引起了人们的关注。膳食多糖可以延缓碳水化合物转化为葡萄糖的过程,抑制消化酶的活性,调节多种信号通路,从而有效降低血糖。然而,多糖的结构与降糖活性之间的关系尚不清楚,这可能限制其开发和应用。
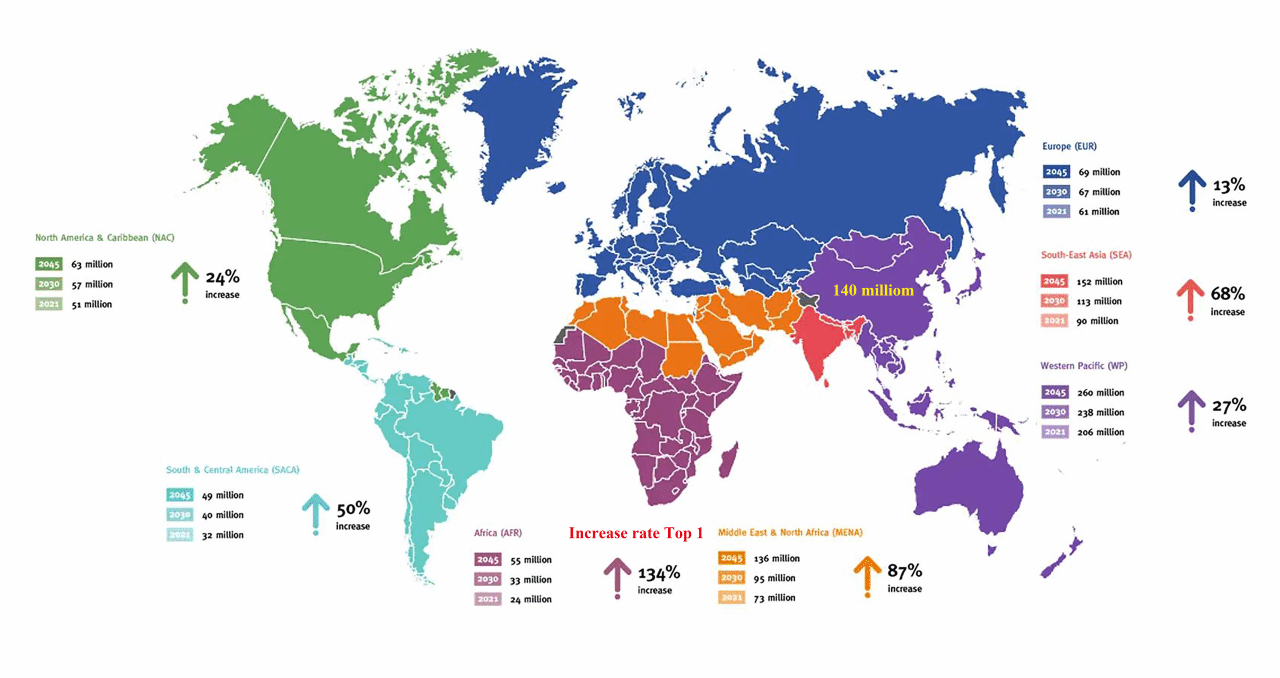
图1 2021年国际糖尿病联盟地图
国家食用菌加工技术研发专业中心康文艺教授、陈晓宇博士等,在本文中聚焦食源性多糖的降糖活性,总结了具有糖代谢调节作用的多糖的来源、结构特征(单糖组成、分子量、构象等),讨论了其构效关系,并从消化酶、糖代谢、脂代谢、炎症、氧化应激、肠道菌群等方面重点综述了食源性多糖调节糖代谢的机理。
Results and Discussion
食源性多糖的来源
食源性多糖主要来源于可食用植物、动物、微生物(包括真菌)等。越来越多的报道表明,食用植物和真菌多糖在调节糖代谢中起着至关重要的作用。可食用植物中获取的多糖,包括中药和药食同源植物、谷物、茶、蔬菜、水果、藻类和其他,已被证实在体内和体外具有潜在的降血糖功能(详见原文)。
具有糖代谢调节作用的食源性多糖的特征
分子量
以往的研究表明,生物活性多糖的重均分子量通常为5-30 kDa,具有降糖活性的多糖的分子量通常在3-150kDa之间。由于分子内氢键较弱,分子量较低的多糖将具有更多的游离氨基和羟基,更有利于多糖发挥生物活性。根据前人的报道,文章汇总了118种来自不同植物和真菌等的食源性多糖的结构特征,其中25个多糖的分子量为5 ~ 30 kDa, 23个多糖的分子量为50 ~ 1000 kDa, 21个多糖的分子量为1000 ~ 10, 000 kDa, 2个多糖的分子量为10,000 ~ 100,000 kDa,只有1个多糖的分子量小于5 kDa。表明分子量越低,多糖的降糖活性越强,小分子量的多糖可能能与糖代谢有关的酶或蛋白紧密结合,从而发挥其降糖作用(详见原文)。
单糖组成
根据以往的研究,在调节糖代谢的食源性多糖中经常观察到10种单糖:葡萄糖、半乳糖、阿拉伯糖、甘露糖、鼠李糖、木糖、半乳糖醛酸、葡萄糖醛酸、岩藻糖、果糖(图2)。此外,在具有降糖功能的食源性多糖中发现了核糖(9次)、半乳糖胺(5次)、N-乙酰-D-(+)-葡萄糖糖胺(1次)。葡萄糖、半乳糖和阿拉伯糖的出现频率最高。一些单糖总是同时存在于食源性多糖中,如葡萄糖、半乳糖、阿拉伯糖或葡萄糖、半乳糖、阿拉伯糖、甘露糖和鼠李糖。半乳糖醛酸和葡萄糖醛酸的存在表明,这些多糖中可能含有少量的酸性多糖,半乳糖醛酸含量高于葡萄糖醛酸。多糖的降糖活性与半乳糖、鼠李糖、甘露糖、葡萄糖醛酸和阿拉伯糖的含量有关。在具有降糖活性的多糖中观察到中性糖比例低,葡萄糖醛酸比例高。
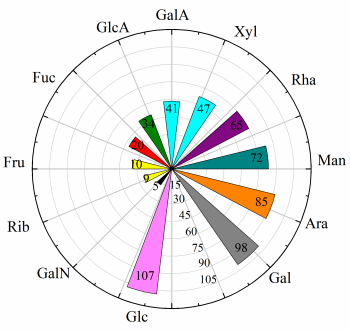
图2 食源性多糖的单糖组成风向玫瑰图
结构特点
在具有降糖潜力的多糖中(表2,详见原文),1→3、1→4和1→6糖苷键总是被观察到,其中1→3糖苷键出现的频率最高。此外,葡聚糖和杂多糖被认为是这些食源性多糖的主要结构类型。根据以往的报道,β-(1→3)-葡聚糖总是出现在食用菌多糖中,约占总数的17.13%,杂多糖占总数的14.81%,葡聚糖和杂葡聚糖的出现频率更高(前5),β-(1→3)-葡聚糖是食用菌多糖的主要结构类型。具有降糖作用的多糖的关键结构单元可能是1→3糖苷键。
食源性多糖的糖代谢调节机制
消化酶
淀粉酶、α-葡萄糖苷酶、β-葡萄糖苷酶、纤维素酶、β-半乳糖糖苷酶等是人体肠道中常见的消化酶,在人体能量代谢中起着至关重要的作用。餐后,由于这些酶的作用,食物总是被转化为葡萄糖和其他物质,进一步为组织细胞提供能量,导致血糖升高。然而,通过抑制其活性,可以有效地抑制餐后血糖的升高。本文总结了多种食源性多糖通过同时抑制α-葡萄糖苷酶和α-淀粉酶活性,仅抑制α-葡萄糖苷酶或α-淀粉酶活性发挥降糖活性的机制。另一方面,食源性多糖形成的溶液粘度和溶液中的网状结构,可能有效地阻止酶(α-淀粉酶等)与淀粉的接触,进一步起到抑制作用(详见原文)。
糖代谢
增强肝脏和外周组织(白色脂肪组织、骨骼肌等)对葡萄糖的摄取,抑制肠道葡萄糖的再吸收,促进机体糖原合成、阻碍糖异生、加速糖酵解过程、促进胰岛素分泌被认为均是降低血糖的有效策略,本文通过文献调研,从葡萄糖摄取、糖原合成、糖异生、糖酵解、胰岛素分泌5个方面详细综述了食源性多糖调节糖代谢的机理(图3,详见原文)。
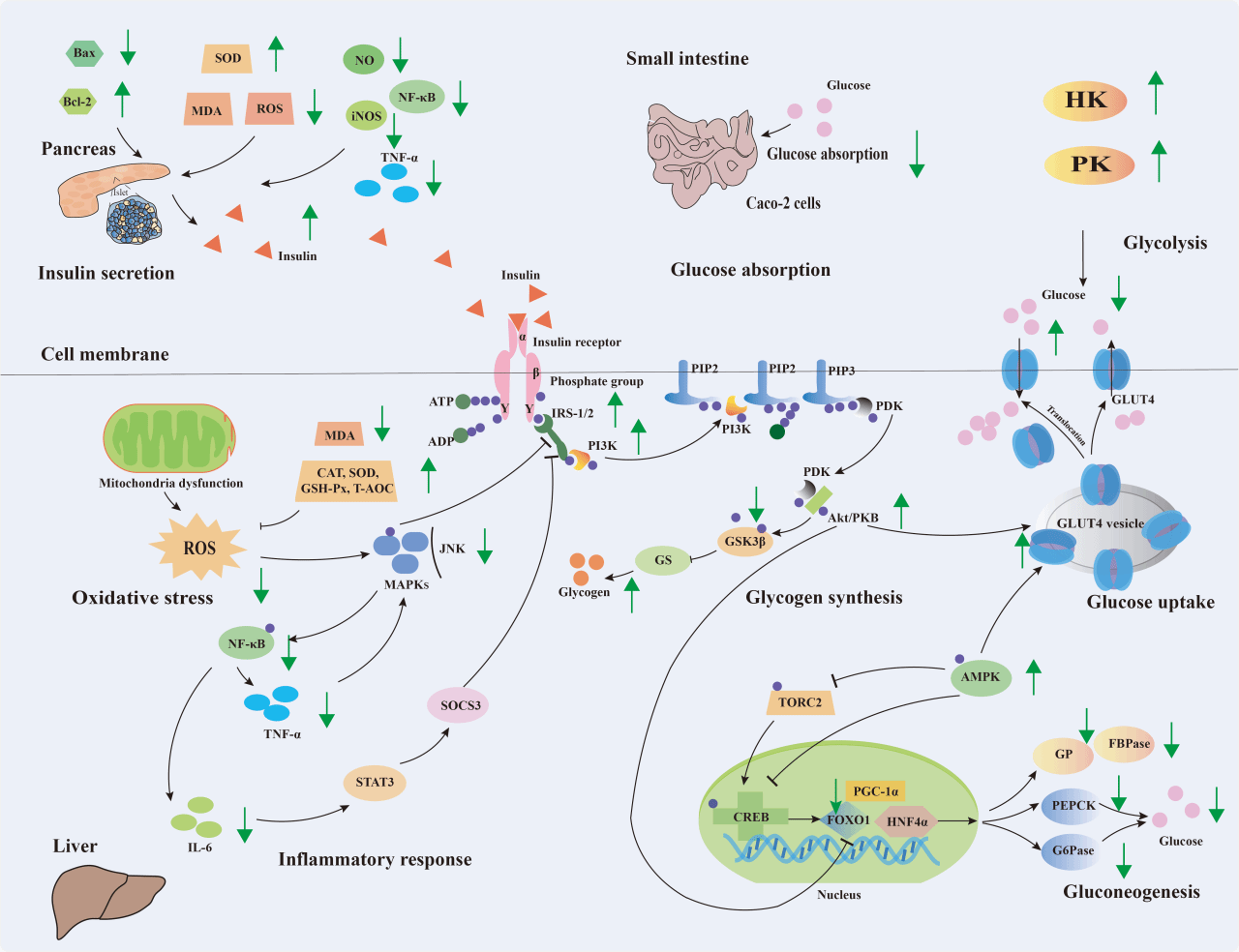
图3 食源性多糖的降糖机制
另外,食源性多糖还可以通过调节脂质代谢、炎症反应和氧化应激等途径间接地参与糖代谢调节,维持葡萄糖稳态(详见原文)。
肠道菌群
越来越多的报道表明,多糖可以增加糖尿病小鼠肠道菌群多样性,通过上调有益菌群(Bacteroides ,Prevotella,Lactobacillus等)丰度,下调有害菌群(Escherichia-Shigella等)丰度,来维持肠道微生态平衡,缓解Ⅱ型糖尿病。多糖的不消化性和胃酸稳定性是其调节肠道菌群和代谢产物的必要条件。靶向肠道菌群也被认为是改善糖代谢紊乱的有效策略。图4显示了食源性多糖基于肠-肝轴的降糖机制(详见原文)。
在消化过程中未被完全消化的大分子多糖可到达肠道进行发酵,从而改变肠道菌群并产生短链脂肪酸等代谢物。这些代谢物可直接或间接进入体循环,激活肠上皮细胞、骨骼肌细胞和肝细胞中的AMPK等,发挥糖代谢调节作用(详见原文)。
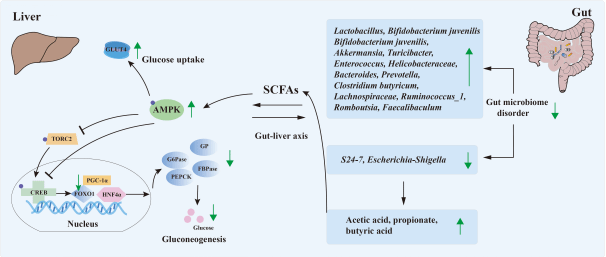
图4 食源性多糖基于肝-肠轴的降糖机制
conclusion
本文综述了具有降糖功能的食源性多糖的研究进展,包括其来源(共60种)、分类(中药、药食同源植物、谷物、茶、蔬菜、水果、藻类、药食同源真菌等)、特征(结构类型:葡聚糖和杂多糖;分子量:5-200kDa;单糖组成前5:葡萄糖、半乳糖、阿拉伯糖、甘露糖、鼠李糖;糖苷键:1→3糖苷键等)和调节糖代谢的分子机制,以获得其降糖作用的构效关系,为功能食品的开发提供理论依据。未来借助创新的技术和手段,进一步揭示这些食源性多糖的结构规律,推动其构效关系的解析,有望为多糖结构及其他天然产物的研究提供新的启示。
原文网址:https://doi.org/10.1016/j.fbio.2024.105679